Melhorar as mitocôndrias e a estabilidade do RE ajuda a eliminar a degeneração do neurônio motor superior que ocorre devido à toxicidade do mSOD1 e à patologia do TDP-43
Barış Genç et al. Clin Transl Med. 2021 fevereiro.
Resumo
Antecedentes: Os neurônios motores superiores (UMNs) são um componente chave do circuito dos neurônios motores. Sua degeneração é uma marca registrada de doenças, como paraplegia espástica hereditária (PHS), esclerose lateral primária (ELP) e esclerose lateral amiotrófica (ELA). Atualmente não existem ensaios pré-clínicos que investiguem as respostas celulares dos UMNs ao tratamento composto, mesmo para doenças dos UMNs. A base da vulnerabilidade dos UMN não é totalmente compreendida e ainda não foi identificado nenhum composto para melhorar a saúde dos UMN doentes:dois grandes obstáculos à construção de estratégias de tratamento eficazes.
Métodos: Novos modelos de repórter UMN, nos quais os UMNs que estão doentes devido à toxicidade da proteína superóxido dismutase mal dobrada (mSOD1) e à patologia TDP-43 são marcados com expressão de eGFP, permitem a avaliação direta da resposta do UMN ao tratamento composto. A microscopia eletrônica revela aspectos muito precisos do retículo endoplasmático (RE) e do dano mitocondrial. A administração de NU-9, um composto inicialmente identificado com base na sua capacidade de reduzir a toxicidade do mSOD1, tem um impacto profundo na melhoria da saúde e da estabilidade dos UMNs, conforme identificado por análises celulares e ultraestruturais detalhadas.
Resultados: Problemas com mitocôndrias e RE são conservados em UMNs doentes entre diferentes espécies. NU-9 tem propriedades farmacocinéticas semelhantes às de drogas. Não possui toxicidade e atravessa a barreira hematoencefálica. NU-9 melhora a integridade estrutural das mitocôndrias e ER, reduz os níveis de mSOD1, estabiliza os dendritos apicais de UMN em degeneração, melhora o comportamento motor medido pelo teste do fio pendurado e elimina a degeneração contínua de UMNs que adoecem tanto por causa da toxicidade de mSOD1 quanto pela patologia TDP-43, duas causas abrangentes distintas e importantes de degeneração de neurônios motores.
Conclusões: As abordagens de descoberta de medicamentos focadas em mecanismos e baseadas em células não apenas abordaram os principais defeitos celulares responsáveis pela perda de UMN, mas também identificaram NU-9, o primeiro composto a melhorar a saúde de UMNs doentes, neurônios que degeneram em pacientes com ELA, HSP, PLS e ELA/FTLD.
Palavras-chave: ELA; PAS; NU-9; POR FAVOR; Patologia TDP-43; mSOD1; neurônios motores superiores.
© 2021 Os Autores. Medicina Clínica e Translacional publicada por John Wiley &Sons Australia, Ltd em nome do Instituto de Bioinformática Clínica de Xangai.
Isenção de responsabilidade do PubMed
Declaração de conflito de interesses
Os autores declaram não haver conflito de interesses.
Números
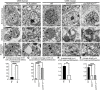 FIGURA 1
FIGURA 1 Os neurônios motores superiores (UMNs) apresentam defeitos ultraestruturais em pacientes com esclerose lateral amiotrófica (ELA) e em modelos de camundongos que estão doentes devido a diferentes causas subjacentes. (A) Imagem representativa de microscopia eletrônica (EM) de UMN de controle normal aparece intacta enquanto (B) UMN de paciente com ELA apresentando defeitos citoarquitetônicos. (C) imagem EM de UMN de mouse WT. (D) Imagem EM representativa de UMN de hSOD1G93A e (E) camundongo prpTDP-43A315T exibindo desintegração ultraestrutural maciça. (F) As mitocôndrias em um controle normal mostrando membranas mitocondriais internas intactas (setas), em oposição a (G) mitocôndrias em pacientes com ELA que apresentam desintegração da membrana mitocondrial interna (cabeças de setas). (H) As mitocôndrias em camundongos WT parecem estar estruturalmente intactas com membranas mitocondriais internas e externas distintas (seta), enquanto (I) mitocôndrias em UMN em um camundongo hSOD1G93A e (J) prpTDP-43A315T exibindo desintegração severa das membranas mitocondriais internas (cabeças de setas). (K) Micrografias eletrônicas do retículo endoplasmático UMN (ER) em um controle normal exibem cisternas longas adequadamente empilhadas (setas), mas (L) ER em paciente com ELA mostra distensão e balão das cisternas do RE (cabeças de setas). Da mesma forma, (M) ER em um camundongo WT (setas) parece estruturalmente intacto em contraste com (N) ER em UMN de hSOD1G93A e (O) camundongo prpTDP-43A315T que exibe cisternas de ER quebradas, curtas e desintegradas (cabeças de setas). (P) Quantificação da porcentagem média de ER com defeitos citoarquitetônicos/UMN em pacientes com ELA. ****p < .0001, teste t de Student. (Q) Quantificação da porcentagem média de ER com defeitos citoarquitetônicos/UMN em hSOD1G93A. ****p < .0001 e camundongos prpTDP-43A315T. ****p < .0001, ANOVA unidirecional seguida pelo teste post hoc de comparações múltiplas de Tukey. (R) Quantificação do comprimento médio das cisternas do RE/UMN em pacientes com ELA. **p < .001, teste t de Student. (S) Quantificação do comprimento médio das cisternas ER/UMN em hSOD1G93A **** p < 0,0001 e camundongos prpTDP-43A315T. ****p < .0001, ANOVA unidirecional seguida pelo teste post hoc de comparações múltiplas de Tukey. Barras de escala:A–E = 2 µm; F–O = 200 nm
 FIGURA 2
FIGURA 2 O tratamento com NU-9 melhora a integridade ultraestrutural das mitocôndrias e do retículo endoplasmático (ER) dos neurônios motores superiores (UMNs) que adoecem devido à toxicidade do mutante SOD1. (A) Progressão do acerto para levar ao NU-9 com estrutura química do NU-9. (B) Desenho experimental para estudos in vivo. (C – F) Imagens representativas de microscopia eletrônica (EM) de UMNs no córtex motor de camundongos WT-UeGFP tratados com veículo em P120. As setas apontam para mitocôndrias com membrana interna intacta e RE com cisternas intactas. Barras de escala, C:2 µm; D–F:200 nm. (G – J) Imagens EM representativas de UMNs no córtex motor de camundongos hSOD1G93A-UeGFP tratados com veículo em P120. (G) O citoplasma é praticamente desprovido de organelas-chave importantes e (H) existem numerosos agregados elétron-densos (pontas de seta) e gotículas grandes. (I) Mitocôndrias que perderam a integridade de sua membrana interna (ponta de seta) ou apresentam danos estruturais gerais e (J) pedaços fragmentados do RE (pontas de seta) são evidentes. Barras de escala, G:2 µm; H-J:200 nm. (K – N) Imagens EM representativas de UMNs no córtex motor de camundongos hSOD1G93A-UeGFP tratados com 100 mg/kg/dia de NU-9. (K) Melhora geral do citoplasma com presença de numerosas organelas. (L) As mitocôndrias parecem saudáveis (setas) e o citoplasma carece de grandes agregados densos e gotículas. (M) A integridade da membrana mitocondrial interna e da estrutura das cristas é restaurada (setas), e (N) as cisternas do ER (setas) estão dispostas em estrutura adequada, sem qualquer fragmentação. Barras de escala, I:2 µm; L–N:200 nm. (O) Quantificação do número total de mitocôndrias/UMN em camundongos hSOD1G93A-UeGFP com tratamento NU-9. **p < 0,006, ANOVA unidirecional seguida pelo teste post hoc de comparações múltiplas de Tukey. (P) Quantificação da porcentagem de mitocôndrias/UMN saudáveis em camundongos hSOD1G93A-UeGFP com tratamento NU-9. ****p < .0001, ANOVA unidirecional seguida pelo teste post hoc de comparações múltiplas de Tukey. (Q) Quantificação do número de cisternas ER/UMN em camundongos hSOD1G93A-UeGFP com tratamento NU-9. *p < 0,016, ANOVA unidirecional seguida pelo teste post hoc de comparações múltiplas de Tukey. (R) Quantificação do comprimento médio das cisternas ER/UMN em camundongos hSOD1G93A-UeGFP com tratamento NU-9. **p < 0,002, ANOVA unidirecional seguida pelo teste post hoc de comparação múltipla de Tukey
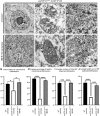 FIGURA 3
FIGURA 3 O tratamento com NU-9 reduz os níveis de SOD1 mal dobrados em neurônios motores superiores (UMNs) de camundongos hSOD1G93A-UeGFP. (A) Imagens representativas de UMNs e coloração de anticorpo B8H10 que reconhece a proteína SOD1 mal dobrada no córtex motor de WT-UeGFP ou (B) camundongos hSOD1G93A-UeGFP tratados com veículo, (C) 20 mg/kg/dia NU-9, ou (D) 100 mg/kg/dia NU-9. Barras de escala, 20 µm; n ≥ 3 réplicas biológicas. (E) Densidade integrada média de fluorescência de SOD1 mal dobrada em UMNs no córtex motor de camundongos WT-UeGFP ou hSOD1G93A-UeGFP tratados com veículo, 20 mg/kg/dia NU-9 ou 100 mg/kg/dia NU-9; média, SEM e pontos de dados individuais mostrados para n ≥ 3 réplicas biológicas. **p < .01, ****p < .0001, ANOVA unidirecional seguida pelo teste de comparação múltipla post hoc de Tukey
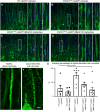 FIGURA 4
FIGURA 4 O tratamento com NU-9 melhora a integridade citoarquitetônica dos dendritos apicais em desintegração dos neurônios motores superiores (UMNs) que ficam doentes devido à toxicidade da SOD1 mal dobrada. (A) Imagens representativas de dendritos apicais UMN no córtex motor de camundongos WT-UeGFP ou (B) hSOD1G93A-UeGFP tratados com veículo, (C) 20 mg/kg/dia NU-9 ou (D) 100 mg/kg/dia NU-9. As áreas em caixa são ampliadas à direita e exemplos adicionais são fornecidos. Barras de escala:50 µm (mag baixa), 10 µm (inserção de alta mag); n ≥ 6 réplicas biológicas. (E) imagem representativa de um dendrito apical saudável, intacto e (F) doente e em desintegração. Barras de escala:5 µm. (G) Porcentagem média de dendritos apicais UMN com vacúolos por seção no córtex motor; média, SEM e pontos de dados individuais mostrados para n ≥ 6 réplicas biológicas. *p < 0,05, **p < 0,01, ANOVA unidirecional seguida pelo teste de comparação múltipla post hoc de Tukey
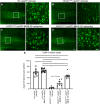 FIGURA 5
FIGURA 5 O tratamento com NU-9 reduz a degeneração do neurônio motor superior (UMN) de UMNs doentes resultante da toxicidade de SOD1 mal dobrada in vivo. (A – D) Imagens representativas de UMNs no córtex motor de camundongos WT-UeGFP ou hSOD1G93A-UeGFP tratados com veículo, 20 mg/kg/dia NU-9 ou 100 mg/kg/dia NU-9. Barras de escala:50 µm; n ≥ 5 réplicas biológicas. (E) Número médio de UMNs por seção no córtex motor; média, SEM e pontos de dados individuais mostrados para n ≥ 5 réplicas biológicas. *p < 0,05, ***p < 0,001, ****p < 0,0001, ANOVA unidirecional seguida pelo teste de comparação múltipla post hoc de Tukey
FIGURA 6 O tratamento com NU-9 melhora a integridade ultraestrutural das mitocôndrias e do retículo endoplasmático (RE) dos neurônios motores superiores (UMNs) que adoecem devido à patologia TDP-43. Defeitos mitocondriais e ER nos UMNs de camundongos prpTDP-43A315T-UeGFP foram publicados anteriormente. (A – C) Imagens representativas de microscopia eletrônica de UMNs de camundongos prpTDP-43A315T-UeGFP não tratados. (A) soma UMN, com poucas organelas intactas. (B) As mitocôndrias perdem a integridade de sua membrana interna (pontas de setas) e (C) as cisternas do RE são desintegradas e quebradas (pontas de setas). (D-F) Imagens representativas de microscopia eletrônica de UMNs de camundongos prpTDP-43A315T-UeGFP tratados com 100 mg/kg/dia de NU-9. (D) Uma melhoria geral na citoarquitetura é evidenciada com membrana nuclear adequada, presença de numerosas organelas saudáveis e falta de agregados elétron-densos. (E) As mitocôndrias parecem saudáveis com membrana interna e cristas melhoradas (seta) (F), e cisternas do RE dispostas em estrutura adequada com ribossomos anexados (setas). Barras de escala:A,D:2 µm; B,C,E,F:200 nm. (G) Quantificação do número total de mitocôndrias/UMN em camundongos prpTDP-43A315T-UeGFP com 100 mg/kg/dia de tratamento NU-9; *p < .03. (H) Quantificação da porcentagem de mitocôndrias/UMN saudáveis em camundongos prpTDP-43A315T-UeGFP com 100 mg/kg/dia de tratamento NU-9; ****p < .0001. (I) Quantificação do número de cisternas ER/UMN em camundongos prpTDP-43A315T-UeGFP com 100 mg/kg/dia de tratamento NU-9; ***p < .0003. (J) Quantificação do comprimento médio das cisternas ER/UMN em camundongos prpTDP-43A315T-UeGFP com 100 mg/kg/dia de tratamento NU-9; ****p < .0001. ANOVA unidirecional seguida pelo teste de comparação múltipla post hoc de Tukey foi usada para análises estatísticas
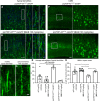 FIGURA 7
FIGURA 7 O tratamento com NU-9 melhora a integridade citoarquitetural dos dendritos apicais em desintegração e elimina a degeneração progressiva dos neurônios motores superiores (UMNs) que ficam doentes devido à patologia TDP-43. (A) Imagens representativas de dendritos apicais UMN no córtex motor de camundongos prpTDP-43A315T-UeGFP não tratados e (B) camundongos prpTDP-43A315T-UeGFP tratados com 100 mg/kg/dia de NU-9. A área em caixa é ampliada à direita com exemplos representativos adicionais. Barras de escala:50 µm (mag baixa), 10 µm (inserção de alta mag); n ≥ 3 réplicas biológicas. (C) imagem representativa de um dendrito apical saudável, intacto e (D) doente e em desintegração. Barras de escala:5 µm. (E) Porcentagem média de dendritos apicais com vacúolos por área de secção no córtex motor; média, SEM e pontos de dados individuais mostrados para n ≥ 3 réplicas biológicas. **p < 0,01, ***p < 0,001, ANOVA unidirecional seguida de teste post hoc de comparações múltiplas de Tukey. (F) Imagens representativas de UMNs no córtex motor de camundongos prpTDP-43A315T-UeGFP não tratados e (G) prpTDP-43A315T tratados com 100 mg/kg/dia de NU-9. Barras de escala:50 µm; n ≥ 3 réplicas biológicas. (H) Número médio de UMNs por área de seção no córtex motor; média, SEM e pontos de dados individuais mostrados para n ≥ 3 réplicas biológicas. *p < .05, **p < .01, ***p < .001, ANOVA unidirecional seguida pelo teste de comparação múltipla post hoc de Tukey
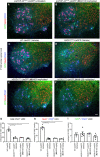 FIGURA 8
FIGURA 8 O tratamento com NU-9 não melhora a degeneração do neurônio motor inferior (LMN) resultante da toxicidade mal dobrada da SOD1 in vivo. (A e B) Imagens representativas de LMNs na medula espinhal lombar de camundongos prpTDP-43A315T-UeGFP não tratados ou tratados com 100 mg/kg/dia de NU-9. Barra de escala:50 µm; n ≥ 3 réplicas biológicas. (C – F) Imagens representativas de LMNs na medula espinhal lombar de camundongos WT-UeGFP ou hSOD1G93A-UeGFP tratados com veículo, 20 mg/kg/dia NU-9 ou 100 mg/kg/dia NU-9. Barra de escala:50 µm; n = 5 réplicas biológicas. (G) Número médio de LMNs ChAT+ por seção na medula espinhal lombar; média, SEM e pontos de dados individuais mostrados para n = 5 réplicas biológicas. ****p < .0001, ANOVA unidirecional seguida pelo teste post hoc de comparações múltiplas de Tukey. (H) Número médio de LMNs NeuN+/ChAT+ (vulneráveis à degeneração) por seção na medula espinhal lombar; média, SEM e pontos de dados individuais mostrados para n = 5 réplicas biológicas. ****p < .0001, ANOVA unidirecional seguida pelo teste post hoc de comparações múltiplas de Tukey. (I) Número médio de LMNs GFP+/ChAT+ (resistentes à degeneração) por seção na medula espinhal lombar; média, SEM e pontos de dados individuais mostrados para n = 5 réplicas biológicas
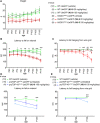 FIGURA 9
FIGURA 9 Dados comportamentais de camundongos WT-UeGFP, hSOD1G93A-UeGFP e prpTDP-43A315T-UeGFP com ou sem tratamento com NU-9. Camundongos WT-UeGFP e hSOD1G93A-UeGFP foram testados a cada 7 dias entre o dia pós-natal (P)60 e P120; Camundongos prpTDP-43A315T-UeGFP foram testados em P60, P90 e P120. (A) Peso dos ratos em gramas. Média e SEM mostradas para n ≥ 5 ratos por grupo. WT-UeGFP (veículo) versus hSOD1G93A-UeGFP (veículo); #p < 0,05, ANOVA bidirecional com teste de comparações múltiplas de Tukey. (B) Latência para cair na aceleração do rotarod em segundos. Média e SEM mostradas para n ≥ 5 ratos por grupo. WT-UeGFP (veículo) versus hSOD1G93A-UeGFP (veículo). #p < .05, ##p < .01, ###p < .001, ####p < .0001, ANOVA bidirecional com teste de comparações múltiplas de Tukey. (C) Latência para cair pendurado de cabeça para baixo em uma grade de arame em segundos. Média e SEM mostradas para n ≥ 5 ratos por grupo. WT-UeGFP (veículo) versus hSOD1G93A-UeGFP (veículo). #p < .05, ###p < .001, ####p < .0001; hSOD1G93A-UeGFP (veículo) versus hSOD1G93A-UeGFP (NU-9 100 mg/kg/dia). *p < 0,05, ***p < 0,001, ****p < 0,0001, ANOVA bidirecional com teste de comparações múltiplas de Tukey. (D) Latência para cair na aceleração do rotarod em segundos. Média e SEM mostradas para n ≥ 4 ratos por grupo. WT-UeGFP (veículo) versus prpTDP-43A315T-UeGFP (veículo). ##p < .01, ####p < .0001, Análise de efeitos mistos com teste de comparação múltipla de Tukey. (E) Latência para cair pendurado de cabeça para baixo em uma grade de arame em segundos. Média e SEM mostradas para n ≥ 3 ratos por grupo. WT‐UeGFP (veículo) versus prpTDP‐43A315T‐UeGFP (veículo), ####p < .0001; prpTDP‐43A315T‐UeGFP (não tratado) versus prpTDP‐43A315T‐UeGFP (NU‐9 100 mg/kg/dia), ****p < 0,0001, análise de efeitos mistos com teste de comparação múltipla de Tukey
Referências
-
- Jara JH, Genc B, Klessner JL, Ozdinler PH. Rotulagem retrógrada, transdução e direcionamento genético permitem a análise celular de neurônios motores corticospinais:implicações na saúde e na doença. Neuroanat frontal. 2014;8:16. - PMC - PubMed
-
- Brunet A, Stuart-Lopez G, Burg T, Scekic-Zahirovic J, Rouaux C. Disfunção do circuito cortical como potencial condutor da esclerose lateral amiotrófica. Neurosci frontal. 2020;14:363. - PMC - PubMed
-
- McColgan P, Joubert J, Tabrizi SJ, Rees G. O microcircuito do córtex motor humano:insights para doenças neurodegenerativas. Nat Rev Neurosci. 2020;21(8):401–415. -PubMed
-
- Gunes ZI, Kan VWY, Ye X, Liebscher S. Complexidade emocionante:o papel dos elementos do circuito motor na fisiopatologia da ELA. Neurosci frontal. 2020;14(573):573. - PMC - PubMed
-
- Oudega M, Perez MA. Reorganização corticoespinhal após lesão medular. J Fisiol. 2012;590(16):3647–3663. - PMC - PubMed
Tipos de publicação
Termos MeSH
Substâncias
Subsídios e financiamento